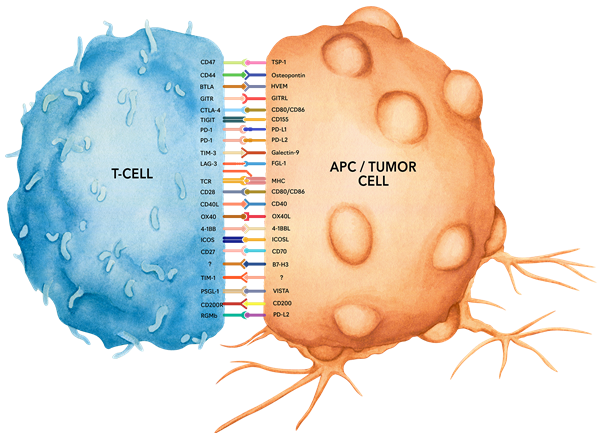
免疫检查点蛋白(Immune Checkpoint Proteins)是一类在免疫系统中起到关键调控作用的分子,它们如同“守门员”一样,能够调节免疫细胞的激活、增殖和功能。这些蛋白通过与相应的配体结合,传递抑制性或刺激性信号,从而维持免疫反应的平衡,防止过度的免疫反应导致自身免疫疾病,同时避免免疫逃逸导致肿瘤的发生和发展。例如,PD-1和CTLA-4是两种典型的抑制性免疫检查点蛋白,它们通过与配体结合,抑制T细胞的过度激活,从而防止自身免疫反应。
如果你想阻断启动免疫检查点途径的特定配体-受体相互作用,你将需要可以依赖的抗体——特别是如果你使用的是体内模型。BioXCell提供广泛的免疫检查点阻断抗体选择,包括抗PD-1,抗PD-L1和抗CTLA-4。BioXCell的抗体纯度高,内毒素含量极低,不含防腐剂或稳定剂,是高灵敏度体内实验的理想选择。
1. CD47 (IAP): CD47广泛表达于造血细胞(例如T和B淋巴细胞、单核细胞、血小板和红细胞)和非造血细胞。CD47参与一系列细胞过程,包括细胞凋亡、增殖、粘附和迁移。此外,它在免疫和血管生成反应中起着关键作用。CD47是血小板反应蛋白-1 (TSP-1) 的受体,TSP-1是一种分泌性糖蛋白,在血管发育和血管生成中发挥作用。已发现CD47在许多不同的肿瘤细胞中过度表达。因此,抗CD47单克隆抗体已被提出并研究作为人类癌症的治疗方法。
货号 | 名称 | 克隆号 | 应用 |
BE0283 | InVivoMAb anti-mouse/human/rat CD47 (IAP) | MIAP410 | 体内封闭CD47 |
BE0270 | InVivoMAb anti-mouse CD47 (IAP) | MIAP301 | 体内封闭CD47 |
2. CD44:小鼠和人CD44 也称为 Hermes、HCAM 和 Pgp-1。CD44 是一种 80-95 kDa 糖蛋白,在所有白细胞、内皮细胞、肝细胞和间充质细胞上表达。作为粘附分子,CD44 参与多种细胞功能,包括淋巴细胞活化、再循环和归巢以及造血。CD44 是透明质酸的受体,还可以与其他配体相互作用,例如骨桥蛋白、胶原蛋白和基质金属蛋白酶 (MMP)。此外,CD44 参与肿瘤转移,抗体靶向 CD44 已被证明可以降低各种肿瘤的恶性活动。有趣的是,白血病细胞上高水平的粘附分子 CD44 对产生白血病至关重要。
货号 | 名称 | 克隆号 | 应用 |
BE0039 | InVivoMAb anti-mouse/human CD44 | IM7 | 体内中和CD44 |
3. BTLA :BTLA是免疫球蛋白超家族(Ig superfamily)的成员,表达于B细胞、T细胞、巨噬细胞、树突状细胞、NK细胞和NKT细胞。与PD-1和CTLA-4类似,BTLA与B7家族的同源蛋白B7-H4相互作用。然而,与PD-1和CTLA-4不同,BTLA通过与肿瘤坏死因子家族受体的相互作用来抑制T细胞活性,而不仅仅是与B7家族的细胞表面受体相互作用。BTLA是疱疹病毒进入介导因子(HVEM)的配体。研究表明,BTLA-HVEM复合物能够负向调节T细胞免疫反应。
货号 | 名称 | 克隆号 | 应用 |
BE0132 | InVivoMAb anti-mouse BTLA (CD272) | 6A6 | 体内刺激BTLA;体内封闭BTLA |
BE0153 | InVivoMAb anti-mouse BTLA (CD272) | PK18.6 | 体内&体外刺激BTLA |
BE0196 | InVivoMAb anti-mouse BTLA (CD272) | PJ196 | 体内封闭BTLA ;体外激活T细胞 |
4. GITR:GITR(糖皮质激素诱导的肿瘤坏死因子受体)在静息T细胞上低水平表达,而在调节性T细胞(Tregs)上高水平表达。在激活的T细胞上,GITR的表达上调,并为T细胞提供共刺激信号。GITR配体(GITRL)存在于B细胞、巨噬细胞、树突状细胞和内皮细胞上,并被认为在调节先天免疫和适应性免疫反应中发挥重要作用。此外,GITR被认为在调节性T细胞维持的免疫优势性自身耐受中发挥关键作用。小鼠基因敲除研究表明,该受体在调节CD3介导的T细胞激活和程序性细胞死亡中具有重要作用。
货号 | 名称 | 克隆号 | 应用 |
BE0063 | InVivoMAb anti-mouse GITR | DTA-1 | 体内刺激GITR |
5. CTLA-4:CTLA-4在活化的T和B淋巴细胞上表达。CTLA-4在结构上类似于T细胞共刺激蛋白CD28,并且两种分子都与 B7家族成员B7-1 (CD80) 和 B7-2 (CD86) 结合。与配体结合后,CTLA-4会负向调节细胞介导的免疫反应。CTLA-4在诱导和/或维持免疫耐受、胸腺细胞发育和调节保护性免疫方面发挥作用。CTLA-4在免疫下调中的关键作用已在CTLA-4缺陷小鼠中得到证实,这些小鼠在 3-5 周龄时因淋巴增生性疾病的发展而死亡。CTLA-4是一组抑制性受体之一,正在通过免疫检查点阻断作为癌症治疗靶点进行探索。
货号 | 名称 | 克隆号 | 应用 |
BE0131 | InVivoMAb anti-mouse CTLA-4 (CD152) | 9H10 | 体内中和CTLA-4 |
InVivoMAb anti-mouse CTLA-4 (CD152) | 9D9 | 体内中和CTLA-4 | |
BE0032 | InVivoMAb anti-mouse CTLA-4 (CD152) | UC10-4F10-11 | 体内中和CTLA-4 |
6. TIGIT:TIGIT是一种26 kDa的I型跨膜蛋白,属于脊髓灰质炎病毒受体(PVR)家族。TIGIT已被发现在小鼠的滤泡T辅助细胞上表达,而在人类中,它在许多T细胞亚群中表达,包括活化T细胞、滤泡T辅助细胞、记忆T细胞和调节性T细胞以及NK细胞。TIGIT可以与PVR和PVR样家族的某些成员相互作用,包括PVR、PVRL2、PVRL3、CD155和CD112。TIGIT被认为可以负向调节NK和T细胞活化。树突状细胞与T细胞上的TIGIT结合导致其分化为耐受性表型,IL-10分泌增加,IL-12产生减少。TIGIT 敲除小鼠更容易患自身免疫性疾病。
货号 | 名称 | 克隆号 | 应用 |
BE0274 | InVivoMAb anti-mouse TIGIT | 1G9 | 体内刺激TIGIT |
7. PD-1:PD-1在CD4和CD8胸腺细胞以及活化的T细胞、B细胞和髓样细胞上短暂表达。在成功清除抗原后,PD-1的表达会下降。此外,在前B细胞阶段发育中的B细胞中也检测到Pdcd1 mRNA的表达。PD-1的结构中包含一个ITIM(基于酪氨酸的免疫受体抑制基序),这表明PD-1可能对T细胞受体(TCR)信号传导具有负向调节作用。PD-1通过与其两个配体PD-L1和PD-L2(均为B7家族成员)结合来传递信号。配体结合后,PD-1的信号传导会抑制T细胞的激活,导致T细胞增殖减少、细胞因子产生降低以及T细胞凋亡增加。此外,PD-1在维持外周耐受和预防自身免疫疾病方面发挥着关键作用,因为PD-1基因敲除的小鼠会出现扩张型心肌病、脾肿大和外周耐受丧失。在许多肿瘤(包括鳞状细胞癌、结肠腺癌和乳腺腺癌)中,PD-L1的诱导表达较为常见。PD-L1的过表达会导致肿瘤细胞对CD8 T细胞介导的溶解作用产生更强的抵抗性。在黑色素瘤小鼠模型中,通过使用阻断PD-L1与其受体PD-1相互作用的抗体治疗,可以暂时抑制肿瘤生长。基于这些原因,目前正在进行抗PD-1免疫疗法作为癌症治疗的研究。
货号 | 名称 | 克隆号 | 应用 |
InVivoMAb anti-mouse PD-1 (CD279) | RMP1-14 | 体内阻断PD-1/PD-L信号 | |
InVivoMAb anti-mouse PD-1 (CD279) | 29F.1A12? | 体内阻断PD-1/PD-L信号 | |
BE0033-2 | InVivoMAb anti-mouse PD-1 (CD279) | J43 | 体内阻断PD-1/PD-L信号 |
8. PD-L1:PD-L1在T细胞、B细胞、NK细胞、树突状细胞以及IFNγ刺激的单核细胞、上皮细胞和内皮细胞上表达。PD-L1与其受体PD-1结合,而PD-1则存在于CD4和CD8胸腺细胞以及激活的T细胞、B细胞和髓系细胞上。PD-L1与PD-1的结合会导致T细胞受体(TCR)介导的T细胞增殖和细胞因子产生的抑制。PD-L1被认为在肿瘤免疫逃逸中发挥重要作用。在许多肿瘤中,诱导PD-L1的表达较为常见,这会导致肿瘤细胞对CD8 T细胞介导的溶解作用产生更强的抵抗性。在黑色素瘤小鼠模型中,通过使用阻断PD-L1与PD-1相互作用的抗体进行治疗,可以暂时抑制肿瘤生长。
货号 | 名称 | 克隆号 | 应用 |
InVivoMAb anti-mouse PD-L1 (B7-H1) | 10F.9G2? | 体内封闭PD-L1 |
9. PD-L2:PD-L2是一种 25 kDa的I型跨膜蛋白,属于免疫球蛋白超家族中的B7家族。PD-L2在单核细胞、巨噬细胞和树突状细胞亚群中表达。PD-L2与其受体PD-1 结合,PD-1存在于CD4和CD8胸腺细胞以及活化的T和B淋巴细胞和髓系细胞中。PD-L2与PD-1结合可抑制TCR介导的T细胞增殖和细胞因子产生。
货号 | 名称 | 克隆号 | 应用 |
BE0112 | InVivoMAb anti-mouse PD-L2 (B7-DC) | TY25 | 体内封闭PD-L2 |
10. TIM-3: TIM-3在Th1淋巴细胞表面特异性高表达,而Th2淋巴细胞表达TIM-1和TIM-2。TIM-3通过与细胞相关C型凝集素半乳糖凝集素9结合而激活。结合后,TIM-3会诱导Th1细胞凋亡。研究表明,抑制小鼠的TIM-3信号传导会加剧实验性自身免疫性脑脊髓炎,促进IFNγ产生和Th1细胞增殖。研究表明,TIM-3也是诱导耐受性所必需的,因为TIM-3基因敲除动物和用TIM-3-Ig融合蛋白治疗的小鼠在诱导抗原特异性耐受性方面均存在缺陷。此外,TIM-3信号传导目前正在被探索作为癌症免疫治疗的靶点,因为表达TIM-3和PD-1的CD8 T细胞在细胞周期进程和效应细胞因子产生方面比单独表达PD-1的细胞表现出更大的缺陷。
货号 | 名称 | 克隆号 | 应用 |
BE0275 | InVivoMAb anti-mouse TIM-3 (CD366) | B8.2C12 | 体内中和TIM-3 |
BE0115 | InVivoMAb anti-mouse TIM-3 (CD366) | RMT3-23 | 体内中和TIM-3 |
11. LAG-3:LAG-3 由活化的T淋巴细胞、NK细胞和T调节细胞表达。LAG-3的主要配体是MHC II类,其结合亲和力甚至高于 CD4。结合后,LAG-3 被认为发挥与 CTLA-4和PD-1类似的作用,包括下调TCR信号传导和抑制CD4依赖性T细胞功能。LAG-3还被证实有助于T调节细胞的抑制功能。与抑制相反,LAG-3已被证实通过激活抗原呈递细胞来促进免疫反应。
货号 | 名称 | 克隆号 | 应用 |
BE0174 | InVivoMAb anti-mouse LAG-3 | C9B7W | 体内中和LAG-3 |
12. 4-1BB (CD137):4-1BB是一种39 kDa的跨膜蛋白,表达于T细胞、NK细胞、树突状细胞、粒细胞和肥大细胞。当其配体4-1BBL结合后,4-1BB通过激活NF-κB、c-Jun和p38下游通路,为CD4和CD8 T细胞提供共刺激信号。4-1BB通路在多种疾病(包括癌症)中的重要性已被多次强调。有研究表明,激动性抗4-1BB抗体能够诱导T细胞介导的抗肿瘤免疫反应。
货号 | 名称 | 克隆号 | 应用 |
BE0169 | InVivoMAb anti-mouse 4-1BB (CD137) | LOB12.3 | 体内激活4-1BB |
BE0239 | InVivoMAb anti-mouse 4-1BB (CD137) | 3H3 | 体内&体外刺激4-1BB |
BE0296 | InVivoMAb anti-mouse 4-1BB (CD137) | 17B5 | 体外封闭4-1BB |
13. 4-1BBL :4-1BBL是一种97 kDa的肿瘤坏死因子(TNF)超家族成员,主要由树突状细胞、巨噬细胞以及活化的B细胞和T细胞表达。4-1BBL与4-1BB(CD137)的相互作用通过激活NF-κB、c-Jun和p38下游通路,为CD4和CD8 T细胞提供共刺激信号。
货号 | 名称 | 克隆号 | 应用 |
BE0110 | InVivoMAb anti-mouse 4-1BBL (CD137L) | TKS-1 | 体内封闭4-1BBL |
更多免疫检查点蛋白体内抗体,详询BioXCell中国代理商艾美捷科技!

产品订购:sales@amyjet.com
邮政编码:430070
公司地址:武汉市洪山区光谷大道35号
光谷总部国际二期时代1栋13楼
提示:本公司所有产品仅供科研使用,不用于临床诊断。
版权所有:艾美捷科技有限公司 鄂ICP备10204150号-1 鄂公网安备:42018502004523号
第二类医疗器械经营备案凭证:鄂汉药监械经营备20234324号

微信扫码在线客服